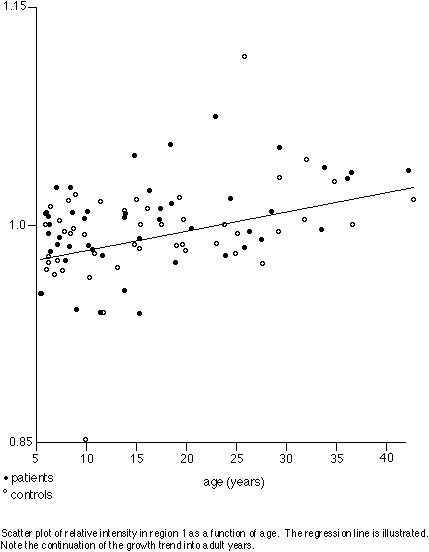
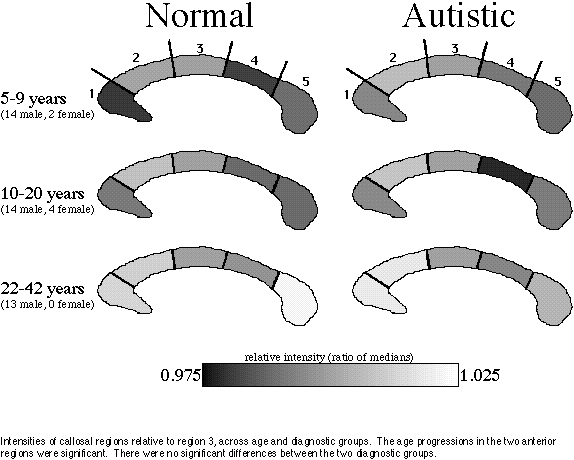
Assuming that callosal intensity increases can be identified with the progress of myelination, our regression results confirm the continuation of anterior callosal myelination into adulthood. Our failure to find any significant intensity differences between patients and controls suggests that the callosal axons that are present in the autistic brain are normally or nearly normally myelinated, and that the age progression of myelination in autistic brains is similar to that in normal brains. Thus our result supports a common mechanism for the parietal sulcal widening and posterior callosal thinning observed in autism: since myelination appears normal, it must be the axons that are absent.
Although no difference was detected between diagnostic groups, there was a significant progression of signal intensity across age groups in the anterior corpus callosum. Fibres of this region originate from lateral prefrontal association cortex, as assayed by studies of Wallerian degeneration in monkeys [Pandya & al. 1971] and by studies of callosal atrophy following focal cortical lesions in humans [de Lacoste & al. 1985]. Continuing development of these fibres may be an anatomical correlate of the final maturation of cognitive abilities and behaviour.
The choice of an internal reference carries benefits and drawbacks. We chose region 3 because it carries mostly fibres from the motor strip [Pandya & al. 1971, de Lacoste & al. 1985] and therefore can be expected to mature earlier than regions carrying fibres from higher-order cortices, and because we have found this region to be unaffected by the callosal narrowing phenomenon of autism [Egaas & al. 1995]. In addition, considering the very slight differences in signal that are being measured, we thought it prudent to give maximum resolution to the ratio by selecting a reference level within or close to the range of the signal being measured. The drawback, of course, is that an increase in signal common to all the comparison regions cannot be distinguished from a decrease in the reference signal. However, we observed no such global perturbation of the measurements; rather, there was a strong, differential increase in the anterior regions.
Of course our failure to detect any difference in callosal intensity between patients and controls does not mean that subtle differences cannot exist. The measurements of this study admit several sources of variance, which we have controlled as tightly as has been practical. Intrinsic variance within the study population can be minimised by using a cross-sequential design [Pujol & al. 1993], a hybrid of cross-sectional and longitudinal designs in which age-related change is assayed by repeating the measurement for each subject. We hope to repeat our scans in order to obtain data for such a design. Signal variance was minimised by our use of the same protocol and the same instrument for all scans, and by our choice of a within-subject reference whose localisation and intensity are similar to that of the region being measured. Finally, variance in morphometric measurements was minimised by use of the median intensity instead of the mean.
Because of the much greater prevalence of autism in males than in females, our patient population and the matched control group were heavily weighted toward males. Thus our results apply primarily to the male brain. Whether sex differences in callosal intensity and cross-sectional area exist remains an open question; sex differences in area have been reported but have not been consistently replicated.
{kind=link}
{kind=link}