Methods
Nineteen subjects participated in the experiment. Four of these were
eliminated from the analysis because of excessive eye movements, and three were
eliminated due to high levels of alpha activity which tended to leak into the
stimulus band and swamp the very small effects being described. Of the
remaining twelve subjects, five were female and seven were male. All were
right-handed, had normal or corrected-to-normal vision, and were between the
ages of 19 and 32.
The paradigm combined elements of the spatial shifting task of Posner &
al. [1984] with the rapid repetition of the intermodality shifting task of
Akshoomoff and Courchesne [1992], and used rapidly flashed stimuli to establish
a steady-state response. Coloured squares, 1.8° on each side, were
displayed at the extreme left and right margins of a video monitor (NEC
MultiSync 3D) so that their centres were 3.0° superior and 5.1° lateral
to a fixation point displayed at the centre of the monitor. The viewing
distance was 59cm. Squares flashed at 8.9s-1 with a 50% duty cycle.
Flashes on the two sides were 180° out of phase with each other, that is,
the offset of a stimulus on one side coincided with the onset of a stimulus on
the other side, and vice versa. Target stimuli were red; non-target
stimuli were green. A spatial cue at the beginning of each trial signalled the
subject to begin the trial with their attention focussed on the cued side and
with a joystick pointed toward the cued side. On detecting a target on the
currently attended side, subjects had as rapidly as possible to shift their
attention to the other side and to move the joystick to the other side.
Stimuli were presented in 144 trials. Each trial contained 32 targets and was
about 50s in duration. Subjects had control of the lengths of rest periods
between trials, and signalled the computer to begin each trial. Before
recording began, subjects practised the task until they felt familiar with it.
Subjects were warned of the possibility of visual fatigue and encouraged to
take liberal breaks between trials. Eye fixation was monitored by
electrooculography and by closed-circuit television. Stimuli were sequenced by
a Markov process such that the pseudorandom intervals between successive
targets on the same side as each other were always greater than 1200ms, the
length of the response window. Joystick movements that followed target stimuli
by 200ms to 1400ms were considered hits. Because of this constraint on
intervals between successive co-lateral targets, joystick movements could be
mapped unambiguously to particular targets.
EEG was recorded using Scan386 digitisation software (Neuroscan, El Paso,
Texas) and a Scientific Solutions analogue-to-digital converter with a Grass
Model 12 Neurodata Acquisition System. Half-amplitude cutoffs were
0.1s-1 and 100s-1.
Ag-AgCl electrodes, 1cm in diameter, were arranged on the occipital scalp in
an 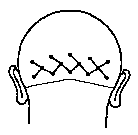 array consisting of three parallel tiers. The lower tier was oriented along
a line across the scalp surface parallel with and 1.5cm superior to a line
passing through the inion and the preauricular point. The other two tiers were
oriented along parallel lines 2cm and 4cm superior to the lower tier. The
lower and upper tiers each contained five electrode sites, placed on the
midline and 4cm and 8cm lateral to the midline on each side. The middle tier
contained four electrode sites, placed 2cm and 6cm lateral to the midline on
each side. The diagonal spacing between electrode centres was thus 2.8cm.
Thirteen bipolar derivations spanned the entire array and recorded information
sufficient for the algebraic derivation of a five-point Hjorth transformation
at each of the four sites in the middle tier. (The Hjorth [1975]
transformation is a discrete approximation to the Laplacian. The use of the
Laplacian emphasises sources that underlie the central electrodes in the array
[Srebro 1985].) This arrangement yielded four virtual channels, one lateral
and one medial on each side of the occipital scalp. Bipolar derivations
between the two outer canthi and from above and below the right eye yielded the
horizontal and vertical electrooculograms. Phase-locked amplitude and
non-phase-locked amplitudes at the 8.9s-1 stimulus delivery
frequency and at its 17.8s-1 harmonic were analysed at each of these
sites using the Gnuroscan SSEP processing system [Belmonte 1997], a
public-domain set of extensions to the commercial Neuroscan software.
array consisting of three parallel tiers. The lower tier was oriented along
a line across the scalp surface parallel with and 1.5cm superior to a line
passing through the inion and the preauricular point. The other two tiers were
oriented along parallel lines 2cm and 4cm superior to the lower tier. The
lower and upper tiers each contained five electrode sites, placed on the
midline and 4cm and 8cm lateral to the midline on each side. The middle tier
contained four electrode sites, placed 2cm and 6cm lateral to the midline on
each side. The diagonal spacing between electrode centres was thus 2.8cm.
Thirteen bipolar derivations spanned the entire array and recorded information
sufficient for the algebraic derivation of a five-point Hjorth transformation
at each of the four sites in the middle tier. (The Hjorth [1975]
transformation is a discrete approximation to the Laplacian. The use of the
Laplacian emphasises sources that underlie the central electrodes in the array
[Srebro 1985].) This arrangement yielded four virtual channels, one lateral
and one medial on each side of the occipital scalp. Bipolar derivations
between the two outer canthi and from above and below the right eye yielded the
horizontal and vertical electrooculograms. Phase-locked amplitude and
non-phase-locked amplitudes at the 8.9s-1 stimulus delivery
frequency and at its 17.8s-1 harmonic were analysed at each of these
sites using the Gnuroscan SSEP processing system [Belmonte 1997], a
public-domain set of extensions to the commercial Neuroscan software.
The vertical electrooculogram was median-filtered in a 75ms window, and the
horizontal electrooculogram was median-filtered in a 336ms window. EEG during
intervals in which change in the filtered vertical electrooculogram over a
period of 300ms exceeded 100µV was marked for rejection due to suspected blink
artefact, and EEG during intervals in which change in the filtered horizontal
electrooculogram over a period of 75ms exceeded 25µV was marked for rejection
due to suspected horizontal saccades. The four subjects mentioned above who
were excluded from the analysis because of excessive saccades were those whose
averaged post-target horizontal electrooculogram still deviated from the
pre-target baseline by 25µV or more even after processing by this rejection
algorithm.
Artefact-free intervals surrounding correctly detected targets were then
averaged by placing the time of target delivery at the zero point of the
averaging epoch. For each target, the epoch extended back in time to the
previous target on the opposite side and forward in time to the next target on
the opposite side. Phase-locked amplitude was calculated by averaging in the
time domain and then computing local amplitude in a 112ms (one stimulus period)
moving window centred on each time point in succession. The local amplitude
computation was implemented by detrending the contents of the window, Fourier
transforming to the frequency domain, and then calculating the length of the
vector formed by the Fourier sine and cosine coefficicients at each frequency
of interest. To preclude an effect of phase on the result of the detrending
operation, only the first and last points in the window were fit; the
intermediate points had no effect on the computation of the linear trend.
Non-phase-locked amplitude was calculated by subtracting the average Fourier
components from the Fourier transform of each single-trial EEG epoch in order
to remove the phase-locked component, calculating the single-trial amplitude
from these adjusted Fourier components, and averaging the resulting
single-trial amplitudes [Mast & Victor 1991].
A variation on this method multiplied the contents of the window by a cosine
bell just before the Fourier transform, in order to minimise spectral leakage.
However, this windowing operation has the drawback of mixing the response at
neighbouring harmonics. Since the detrending operation by itself forces the
endpoints of the window to zero and thus reduces leakage, the data were
examined both with and without the cosine-bell step in order to ascertain its
effects and its side effects.
Amplitudes in response to correctly detected targets were averaged into four
separate groups depending on the amount of time that had elapsed since the
previous correctly detected target. These inter-target interval bins were
56ms-728ms, 840ms-1512ms, 1624ms-2296ms, and 2408ms or longer. These will be
referred to as bins 0, 1, 2, and 3, respectively. Since 2296ms was the longest
possible interval between target presentations, bin 3, unlike the others,
contained only responses that followed at least one missed target. For each of
these time bins, the difference between amplitude in response to left targets
(rightward shifts) and amplitude in response to right targets (leftward shifts)
was computed. Thus a positive deflection indicates an increase in power in
response to left targets or a decrease in power in response to right targets,
whereas a negative deflection indicates an increase in power in response to
right targets or a decrease in power in response to left targets. Each point
was then replaced by the point within a 112ms (one period of the SSVEP) local
neighbourhood whose absolute value was greatest. This transformation has the
effect of stretching the amplitude extrema, and thus increases statistical
power in cases in which the responses of individual subjects are out of phase
with each other. Finally, a pointwise sign test was applied in order to detect
differences in response between the left-target and right-target conditions.
Effects of inter-target interval were detected using Friedman's test across all
levels of intertarget interval. A degenerate case of Friedman's test served as
a post hoc analysis between adjacent levels of intertarget interval
(with twelve subjects in the analysis, the Friedman [[chi]]2
approximation holds even between only two treatments).
Behavioural data were analysed in seven inter-target interval bins,
56ms-392ms, 504ms-728ms, 840ms-1176ms, 1288ms-1512ms, 1624ms-1960ms,
2072ms-2296ms, and 2408ms or longer. For each of these latency bins and for
each target location, both the average response latency for correct detections
and the ratio of correct detections to the total number of detection
opportunities were computed. These were subjected to an analysis of variance
(BMDP program 2V).
Results