Gnuroscan
a set of UNIX-based tools for manipulating NeuroScan data files
This software is distributed under the GNU General Public Licence, version 2. For alternative licensing arrangements, contact the author, Matthew Belmonte.
If you base a publication on this software, please cite either the ACM newsletter publication mentioned in the source files, or (preferably), the first peer-reviewed article in which it was applied (follow the link).
The code described here was developed with a specific application in mind.
However, many of the programs are more generally applicable. Here is a list of
these general-purpose programs that you may find useful:
TRANSFORMATIONS
cntcat - concatenate .CNT files
lderiv - compute a linear derivation from a .CNT file
range - replace points in a .AVG file by local range measures
extreme - replace points in a .AVG file by local extrema
detrend - remove linear voltage trends from a .AVG file (a high-pass filter)
smooth - smooth a .AVG file (a low-pass filter)
rectify - replace points in a .AVG file by their absolute values
power - compute the power spectrum of a .AVG file in a moving window
addavg - compute the pointwise sum of two .AVG files
scale - scale a .AVG file
subepoch - extract a sub-interval from a .AVG file
ztrans - transform a .AVG file into z-scores, with variance and mean from another .AVG file
STATISTICS
chisquare - compute a Chi-square test on the .AVG output from ztrans
permtest - a permutation test applied to a group of .AVG files
ttest - apply Student's t test to a .AVG file
mannwhitney - the Mann-Whitney test applied pointwise to two sets of .AVG files
signtest - the sign test applied pointwise to a group of .AVG files
friedman - Friedman's rank test applied pointwise to a set of .AVG files
DUMPS
dumpavg - create an ASCII dump of a .AVG file
dumpevtb - create an ASCII dump of the event table of a .CNT file
plotavg - translate a .AVG file into PostScript code for printing
Only the "power" program requires the Numerical Recipes library to compile; the others depend only on public-domain libraries.
This paper focusses on the specific application for which Gnuroscan was
developed, the analysis of steady-state evoked potentials. If you just want
the software and aren't interested in steady-state evoked potentials, you may
want to go directly to the download page.
A Software System for Analysis of Steady-State Evoked Potentials
Copyright © 1996 by Matthew Belmonte
Address for correspondence: mkb4@cornell.edu
An abridged version of this paper appears in the Association for Computing Machinery SIGBIO Newsletter, 17:1:9-14 (1997).
ABSTRACT: A suite of computer programs tailored to the analysis of
steady-state evoked potential data in attentional shifting experiments is
described. Stimuli are sequenced pseudorandomly while maintaining constraints
on overlapping of response windows. Averages of phase-locked and
non-phase-locked steady-state amplitudes are computed in variable-length epochs
and corrected for the effect of sampling noise. Various algebraic and
combinational transformations of averaged data are implemented. Several
parametric and nonparametric statistical tests are implemented, and the results
of these procedures are visualised. An application of this software has
revealed modulation of a steady-state visual evoked potential by spatial
attention. The software has been designed with portability in mind, and many
of the programs are applicable to more general problems in evoked potentials.
Descriptors: EEG, Steady-State Evoked Potential, Software
INTRODUCTION
A steady-state evoked potential (SSEP) is a continuously oscillatory
potential generated by the brain in response to a periodic driving stimulus.
Particularly good resonances are observed for the visual SSEP when stimuli are
flashed at about 10s-1 [Fedotchev & al. 1990] and for the
auditory SSEP when clicks occur at about 40s-1 [Galambos &
al. 1981]. Perturbations of the SSEP from its background amplitude and
phase indicate changes in brain state. The SSEP has been used to index the
salience of stimuli in studies of auditory perception [Rohrbaugh & al.
1989; Galambos & Makeig 1992] and visual perception [Allen & al.
1986; Chen & al. 1990; Bach & Meigen 1992; Snowden &
al. 1995], and as an indicator of the degree of involvement of cortex in
cognitive processing tasks [Silberstein & al. 1990, 1995]. The
rapid, regularly timed, and ongoing nature of SSEP resonances makes them useful
in such applications. The temporal resolution of SSEP measurements depends
upon the frequency of the driving input and is bounded by the switching times
of the neural networks involved in generating the SSEP and by the modulation
transfer functions of the overlying tissues and sensors.
In tasks that require some behavioural response, the stimuli that drive the
SSEP may or may not be identical with the stimuli to be evaluated for response.
A target stimulus such as a tone or a picture can perturb an SSEP driven by an
ongoing sequence of background stimuli such as clicks or flashes.
Alternatively, the properties of the background stimuli themselves can be
modulated in order to provide a target stimulus. For example, in an experiment
involving a visual SSEP, a colour change in one of a series of periodic flashes
can indicate a target. Stimuli consist of a series of instances of the
background stimulus punctuated by instances of the target stimulus. The evoked
responses consist not only of the response to the single, target stimulus, but
also of perturbations in the overlapping responses to the surrounding
background stimuli.
 A logical extension of the application of SSEPs as measures of perception
within sensory channels is their use in experiments involving the shifting or
reallocation of attention between sensory channels. These two different
channels might be spatial locations, spatial frequencies, colours, pitches, or
even different sensory modalities. In our test experiment, the two sensory
channels were defined by spatial location. The behavioural task essentially
was two visual oddball paradigms running side by side in two different spatial
locations; a target stimulus in one location cued not only an overt,
behavioural response but also a shift of attention to the other, concurrently
running oddball series. The next target in this newly attended series then
cued a shift back to the original location, and so on. The initial,
immediately practical goal of this project was to design a software system
capable of processing SSEP responses in this paradigm.
A logical extension of the application of SSEPs as measures of perception
within sensory channels is their use in experiments involving the shifting or
reallocation of attention between sensory channels. These two different
channels might be spatial locations, spatial frequencies, colours, pitches, or
even different sensory modalities. In our test experiment, the two sensory
channels were defined by spatial location. The behavioural task essentially
was two visual oddball paradigms running side by side in two different spatial
locations; a target stimulus in one location cued not only an overt,
behavioural response but also a shift of attention to the other, concurrently
running oddball series. The next target in this newly attended series then
cued a shift back to the original location, and so on. The initial,
immediately practical goal of this project was to design a software system
capable of processing SSEP responses in this paradigm.
BACKGROUND
Most conventional software for the processing of evoked potentials is
tailored to a single-stimulus format, in which stimuli are delivered in
isolated instances and the evoked response is a single, self-contained waveform
rather than a perturbation of an ongoing waveform. Also, since target stimuli
generally occur in separate experimental trials from each other, the length of
the averaging epoch in teach trial is unbounded by neighbouring targets, and
therefore an epoch of constant length is assumed. Methods of data reduction
and statistical evaluation usually are not handled by such software systems,
and instead must be specified by the experimenter explicitly. Finally, there
is some difficulty in combining modules in an off-the-shelf manner in order to
automate data processing.
Newer software systems have made some progress against many of the
aforementioned shortcomings. A commercial system in widespread use in evoked
potential laboratories, SCAN (NeuroScan, El Paso, Texas) was used as the basis
for this system. SCAN includes a basic toolkit for the analysis of evoked
potentials, and a batch scripting language that allows automation of many of
the processing steps by modular combination of various subprograms from this
toolkit. Nevertheless, many users of SCAN still employ custom, in-house
software for the less standard and more arcane aspects of their data
processing. In particular, the difficulties of the single-stimulus format, the
constant-length averaging epoch, and the manual nature of data reduction and
statistical evaluation remain to be addressed.
DESIGN CONSIDERATIONS
The goal of the project reported here was to construct an evoked
potential processing system tailored to steady-state applications but also
applicable to other problems in signal processing. Realistic assumptions of
the computing power commonly available in the 1990s were made: the system tends
to trade computational tractability for exactness of results. A benefit of
this trade-off is that few assumptions about the statistical character of the
data need be made; for example, the nonparametric statistical routines produce
valid results even when the data are asymmetrically distributed. There is an
`off-the-shelf' modularity to the system: single operations such as linear
transformation, scoring, averaging, statistical analyses, and visualisation are
implemented as separate programs. This ease of combining the several utilities
in the Gnuroscan toolkit leaves the investigator free to deal with scientific
questions rather than becoming caught up in the specifics of algorithmic
designs and software implementations.
Because I was starting from the basis provided by the SCAN software, the
analysis software described herein, `Gnuroscan', has been implemented to
conform to SCAN's data file formats, but these are easily translated into other
representations. Portability, modularity, and extensibility of the software
were major goals. The entire system has been implemented in ANSI standard C
language, and all graphical output has been implemented using the standard
display language PostScript. Details on the data format, the algorithms, and
the implementation details are available in order to allow extension and
customisation of the software by other investigators.
The software, including
source code, is available from the author.
SYSTEM DESCRIPTION
Sequencing and presentation of stimuli. Before data can be
gathered in such an attentional-shift experiment, the stimuli to be delivered
in each sensory channel (in the case of our test experiment, in each spatial
location) must be sequenced. The position in which a stimulus occurs within a
sequence of stimuli determines the bounds of its response window, that is, the
time interval within which a behavioural response to that stimulus may occur.
A shortcoming of previous experiments involving rapidly repeated shifts of
attention back and forth between two different sensory channels [Akshoomoff
& Courchesne 1992; Courchesne & al. 1994a, 1994b] has been the
difficulty in mapping responses unambiguously to particular stimuli, due to
overlaps of response windows. Typically, a response is scored as a hit or as a
false alarm depending on whether it occurs within the response window following
the delivery of a target stimulus. The interval between the onset of the
target and the opening of this window, [Delta]topen, typically some
200ms, is assumed to be the minimum behavioural reaction time that would be
physiologically possible. The interval between the onset of the target and the
closing of this window, [Delta]tclose, 1400ms in the experiments
cited, is assumed to be the maximum possible latency of a response. However,
an ambiguity ensues if the response windows for two successive targets overlap;
it is not immediately clear which target a particular response should be
associated with. Such conflicts can be resolved using heuristics, for instance
by associating each response with the most recent target within whose window
the response lies.
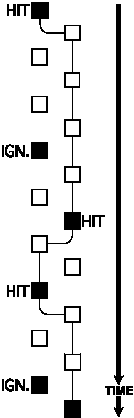
 An ambiguity still ensues, however, in the case in which the response window
for a missed target overlaps the response window for a correctly detected
target in the same sensory channel. In such a situation, the
response cannot be mapped unequivocally to either of the targets. In the
current implementation of the Gnuroscan system, targets alternate between only
two sensory channels. In order to refer to a concrete example, this discussion
will use the case of a spatial attention experiment in which the two channels
correspond to the two sides of space. So, in this example, targets alternate
between the left side and the right side. Therefore, successive targets on the
same side are separated by the sum of two successive intertarget intervals.
Call these intervals ti and ti+1, respectively. Let T be
the time of occurrence of the first target of interest. Then the time of
occurrence of the next target on the same side is
T+ti+ti+1. In order for the response windows not to
overlap, the end of the first window, T+[Delta]tclose, must not be
greater than the beginning of the next window,
T+ti+ti+1+[Delta]topen. This constraint can
be accomplished by generating the sequence of intertarget intervals using a
Markov chain to derive a pseudo-uniform distribution (subject to whatever
discretisation is required by the experimental paradigm or by the stimulus
delivery hardware) of intertarget intervals t over some interval [0,
tmax] conditioned to ti+ti+1 >=
[Delta]tclose-[Delta]topen. This method is applied
off-line, by the sequence program, to compute a stimulus sequence which
is fed to a separate stimulus-delivery program.
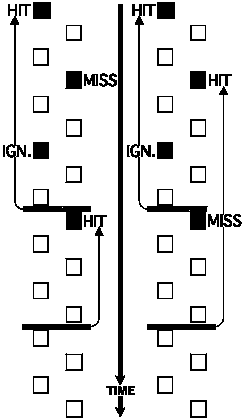
An ambiguity still ensues, however, in the case in which the response window
for a missed target overlaps the response window for a correctly detected
target in the same sensory channel. In such a situation, the
response cannot be mapped unequivocally to either of the targets. In the
current implementation of the Gnuroscan system, targets alternate between only
two sensory channels. In order to refer to a concrete example, this discussion
will use the case of a spatial attention experiment in which the two channels
correspond to the two sides of space. So, in this example, targets alternate
between the left side and the right side. Therefore, successive targets on the
same side are separated by the sum of two successive intertarget intervals.
Call these intervals ti and ti+1, respectively. Let T be
the time of occurrence of the first target of interest. Then the time of
occurrence of the next target on the same side is
T+ti+ti+1. In order for the response windows not to
overlap, the end of the first window, T+[Delta]tclose, must not be
greater than the beginning of the next window,
T+ti+ti+1+[Delta]topen. This constraint can
be accomplished by generating the sequence of intertarget intervals using a
Markov chain to derive a pseudo-uniform distribution (subject to whatever
discretisation is required by the experimental paradigm or by the stimulus
delivery hardware) of intertarget intervals t over some interval [0,
tmax] conditioned to ti+ti+1 >=
[Delta]tclose-[Delta]topen. This method is applied
off-line, by the sequence program, to compute a stimulus sequence which
is fed to a separate stimulus-delivery program.
 Transformation of raw data. Once the raw data have been
collected, the computation begins in earnest. In many analyses, the first step
is to convert the data from the physical recording channels to some linear
combination of the original channels. Examples of this are re-referencing,
transformation between bipolar and common-reference recordings, and Hjorth's
[1975] approximation to the discrete Laplacian transformation. The
cntcat program catenates separate files of continuous EEG data, and the
lderiv program computes an arbitrary linear combination of channels from
a raw data file.
Transformation of raw data. Once the raw data have been
collected, the computation begins in earnest. In many analyses, the first step
is to convert the data from the physical recording channels to some linear
combination of the original channels. Examples of this are re-referencing,
transformation between bipolar and common-reference recordings, and Hjorth's
[1975] approximation to the discrete Laplacian transformation. The
cntcat program catenates separate files of continuous EEG data, and the
lderiv program computes an arbitrary linear combination of channels from
a raw data file.
 Rejection and scoring. The score program assumes that
the horizontal and vertical electrooculograms are available as part of the
recorded electrophysiological data. (These may be virtual channels, derived as
a linear combination of recording channels.) It computes the median of the
horizontal electrooculuogram in a moving window. Because some phasic activity
from the steady-state potential will leak into the eye channels, it's important
that the length of this window be an integral number of interstimulus
intervals. This prevents the biasing of the median by such phasic activity and
thus yields a less variable signal of eye position. When the range of this
signal within a local neighbourhood exceeds a threshold, a horizontal saccade
is detected. In the test experiment, useful values for the length
of the median filter window, the size of the local neighbourhood, and the
rejection threshold were, respectively, 336ms (three complete interstimulus
intervals), 75ms (a liberal figure for the duration of a saccade), and 25uV.
Rejection and scoring. The score program assumes that
the horizontal and vertical electrooculograms are available as part of the
recorded electrophysiological data. (These may be virtual channels, derived as
a linear combination of recording channels.) It computes the median of the
horizontal electrooculuogram in a moving window. Because some phasic activity
from the steady-state potential will leak into the eye channels, it's important
that the length of this window be an integral number of interstimulus
intervals. This prevents the biasing of the median by such phasic activity and
thus yields a less variable signal of eye position. When the range of this
signal within a local neighbourhood exceeds a threshold, a horizontal saccade
is detected. In the test experiment, useful values for the length
of the median filter window, the size of the local neighbourhood, and the
rejection threshold were, respectively, 336ms (three complete interstimulus
intervals), 75ms (a liberal figure for the duration of a saccade), and 25uV.
A similar method is applied to the vertical electrooculogram in order to
detect blinks. Because the amplitude of the blink signal recorded near the
orbit is strictly greater than the amplitude of the SSEP, the length of the
median filter window need not be an integral number of interstimulus intervals
in this case. In the test experiment, useful values for the length of the
median filter window, the size of the local neighbourhood, and the rejection
threshold were, respectively, 75ms, 300ms, and 100uV. Gnuroscan includes an
option for calculating the voltage thresholds automatically from calibrations
marked in the event record.
All this repetitive computation of medians in moving windows is optimised
using a combination of queues and binary search trees. A queue keeps track of
the order of samples within a window, and a binary search tree enables the
implementation of insertion, deletion, and computation of the median in
O(log w) time, where w is the length of the window. Thus, for a
record comprising n digital samples, the entire record can be scanned
for ocular artefacts in O(n log w) time.
The scoring program also assumes that each stimulus and each response are
marked by an event code inserted in the record. Due to the very high rate of
data transfer in SSVEP experiments, with the spacing between event codes
sometimes being as little as a few milliseconds, the software includes checks
for omitted or mis-transcribed event codes. The commercial software used for
data acquisition in our test case did indeed drop some codes, and
mis-transcribed others. These errors probably were due to imperfect
synchronisation of the read operation in the computer that was recording the
data with a strobe signal from the computer that was supplying the event codes.
Whatever the source of the errors, their presence demonstrates the importance
of this check. Isolated corruptions of event codes almost always can be
repaired by utilising redundancies in the data. In our spatial attention
example, suppose the subject is responding to targets in alternate hemifields
by moving a joystick between the two sides of space. In such a situation, it's
impossible for two joystick movements in the same direction to occur without a
joystick movement in the opposite direction between them. For instance, if the
joystick makes a transition from the right side to the left side, it must first
return from the left side to the right side before another transition from
right to left can be recorded. So for each target, if a response occurs within
the current response window, and this response hasn't already been associated
with some prior target, the response is treated as a correct response to the
current target even if its recorded laterality is incorrect. In such a case,
the event code is rewritten with the appropriate laterality. In cases in which
the response code is completely dropped, the response latency cannot be
recovered, but the fact that a response did occur is still evident, because the
omitted response will cause the juxtaposition of two responses that are on the
same side as each other.
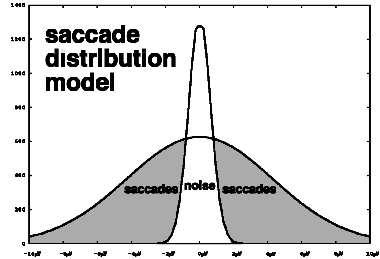 A separate program, eyehand, extracts correlations of horizontal eye
movements with manual responses. The distribution of amplitudes of
maximal-length nondecreasing or nonincreasing intervals of the horizontal
electrooculogram is modelled as the sum of two zero-mean Gaussians, one of them
accounting for noise and microsaccades, and the other accounting for
larger-amplitude saccades of the type that often occur in response to a cue to
shift attention. The Levenberg-Marquardt algorithm [Press &
al. 1992] is used to zero in on the parameters that define these Gaussians.
The absolute value at which these two Gaussian curves cross is computed and
used as a threshold. The saccade latency in response to a target is defined as
the latency of the beginning of the first nondecreasing or nonincreasing
interval whose range exceeds the threshold and whose direction is the same as
the direction of the attentional shift cued by the target. Correlations and
regression coefficients are then computed to describe the relationship between
saccade latency and latency of manual responses.
A separate program, eyehand, extracts correlations of horizontal eye
movements with manual responses. The distribution of amplitudes of
maximal-length nondecreasing or nonincreasing intervals of the horizontal
electrooculogram is modelled as the sum of two zero-mean Gaussians, one of them
accounting for noise and microsaccades, and the other accounting for
larger-amplitude saccades of the type that often occur in response to a cue to
shift attention. The Levenberg-Marquardt algorithm [Press &
al. 1992] is used to zero in on the parameters that define these Gaussians.
The absolute value at which these two Gaussian curves cross is computed and
used as a threshold. The saccade latency in response to a target is defined as
the latency of the beginning of the first nondecreasing or nonincreasing
interval whose range exceeds the threshold and whose direction is the same as
the direction of the attentional shift cued by the target. Correlations and
regression coefficients are then computed to describe the relationship between
saccade latency and latency of manual responses.
 Averaging. The average program is the heart of the
Gnuroscan system. It computes averages both of instantaneous amplitude and of
amplitude within certain frequency bands in a local neighbourhood.
Instantaneous amplitude is calculated by the simple, time-domain averaging
procedure that is widely used in studies of event-related potentials.
Amplitude specific to certain frequency bands is computed by subjecting each
single-trial, time-domain signal to a Fast Fourier Transform (FFT) [Press
& al. 1992], and then calculating the length of the vector formed by
the Fourier sine and cosine coefficients at the frequency or frequencies of
interest. The signal is prepared for the FFT by resampling an interval whose
length is one complete interstimulus period so that the resulting number of
samples within this interval is a power of two, and by detrending and
convolution with a cosine-bell window to prevent spectral leakage. The vector
length of the FFT coefficients is then the amplitude of the signal at the
frequency of interest within a one-stimulus-cycle neighbourhood centred on the
current time point. The FFT interval is stepped through the data in order to
compute successive frequency-specific amplitudes centred on each time point
within the epoch.
Averaging. The average program is the heart of the
Gnuroscan system. It computes averages both of instantaneous amplitude and of
amplitude within certain frequency bands in a local neighbourhood.
Instantaneous amplitude is calculated by the simple, time-domain averaging
procedure that is widely used in studies of event-related potentials.
Amplitude specific to certain frequency bands is computed by subjecting each
single-trial, time-domain signal to a Fast Fourier Transform (FFT) [Press
& al. 1992], and then calculating the length of the vector formed by
the Fourier sine and cosine coefficients at the frequency or frequencies of
interest. The signal is prepared for the FFT by resampling an interval whose
length is one complete interstimulus period so that the resulting number of
samples within this interval is a power of two, and by detrending and
convolution with a cosine-bell window to prevent spectral leakage. The vector
length of the FFT coefficients is then the amplitude of the signal at the
frequency of interest within a one-stimulus-cycle neighbourhood centred on the
current time point. The FFT interval is stepped through the data in order to
compute successive frequency-specific amplitudes centred on each time point
within the epoch.
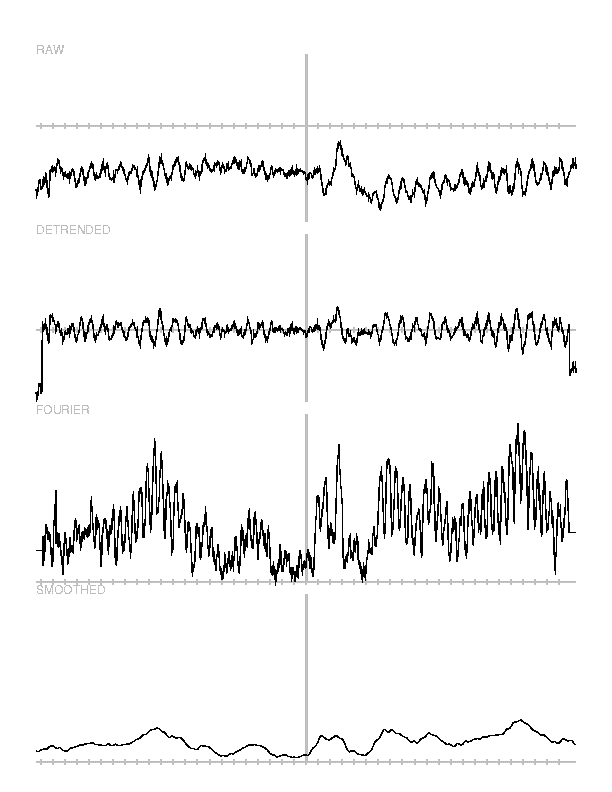
The detrending operation and the windowed amplitude calculation are also
implemented as separate, self-contained programs (detrend and
power), outside of the averaging program. These are used to transform
the instantaneous amplitude averages, which were calculated previously, into
moving-window amplitudes at the frequencies of interest. Why do it both ways?
Why calculate both an average of the single-trial frequency-specific amplitudes
and the frequency-specific amplitudes of a time-domain average? It turns out
that the operations of averaging and of computing the frequency-specific
amplitude are not commutative. That is, one obtains different results when the
amplitude calculation occurs before the averaging than when the averaging
occurs before the amplitude calculation. This is because the amplitude
calculation, depending as it does on a two-dimensional distance relation, is a
non-linear operation. Averaging over sufficiently many trials eliminates (most
of) the variance due to noise and leaves only the variance due to signal.
Noise, in this sense, consists both of instrument noise and of background EEG
whose phase is not time-locked to stimulus presentation. Thus the amplitudes
calculated from the time-domain averages reflect phase-locked activity at each
frequency of interest, whereas the averages of the single-trial amplitudes
reflect total activity, including activity that is not phase-locked. Both
these measures are of interest.
In attempting to maximise signal-to-noise ratio from the data available,
another difficulty arises. In order to squelch as much noise as possible, all
available samples at any particular time point must be included in the
averages. But since the length of time since the previous shift of attention
and the length of time until the next shift of attention vary across different
instances of targets, the epoch length is not constant. The average can still
be computed, by alligning all epochs to the time of target delivery and using a
separate divisor for each time point. But this creates a secondary problem.
The signal-to-noise ratio varies as the square root of the number of samples in
the average. Using this scheme, the number of samples decreases towards the
ends of the maximal-length epoch. So the variance of the signal, and therefore
the amplitude at any particular frequency, is biased upward toward the ends of
the epoch. In pictorial terms, the amplitude function versus time is concave
downward, with a minimum at the centre of the epoch. This bias can be
corrected by using the baseline, that is, the half of the epoch before the time
of target delivery, as a model of the bias. The local range of the signal over
one complete stimulus cycle is fit to the function
(1+tanh(n-p0))/2 * p1/sqrt(p0) +
(1-tanh(n-p0))/2 * p1/sqrt(n)
with parameters p0 and p1 and number of
observations n, using the Levenberg-Marquardt algorithm. In the
expression above, the left-hand factor of each term determines the relative
contribution of the corresponding right-hand factor. The two left-hand factors
are complementary, having unit sum. They are sigmoid functions that implement
a rather abrupt, yet continuous (and so differentiable) switch between
dominance of the left-hand term at high n and dominance of the
right-hand term at low n. When the value of n, the number of
observations in the average, is high, it has little effect on the range of the
signal. Conversely, when n is low, noise biases the amplitude upward
with an inverse dependency on the square root of n. The fitted
parameter p0 is the critical value of n at which the
inverse-square-root, noise-related amplitude level begins to dominate the
constant, asymptotic amplitude level. The noise correction factor for some
particular number of observations nt is then
2/((1+tanh(nt-p0)) +
(1-tanh(nt-p0))*sqrt(p0/nt))
In addition to computing these averages for each combination of target channel,
response accuracy, and length of time since the most recent shift, the
averaging routine calculates the vector of standard deviations associated with
each average, reports average response latency and its standard deviation for
each sensory channel and for several separate ranges of inter-target intervals,
and computes histograms of the amplitude of the horizontal electrooculogram
versus time during attentional shifts.
Transformations of averaged files. Several programs are
provided in order to implement algebraic and combinational operations on
averaged data files. The most basic of these are addavg and
scale. addavg takes a list of averaged files and produces an
averaged file that is the pointwise sum of all these inputs. scale
takes a scalar value and an averaged file and produces the averaged file that
is the product of these two inputs. scale can be applied with a
negative scalar and followed by addavg in order to implement
substraction. scale can be applied with a reciprocal scalar and
preceded by addavg in order to construct a grand average.
The subepoch program truncates an averaged data file at either end.
This is useful in situations in which the epoch includes data points that are
unused in the current analysis. The smooth program replaces each data
point by the average of all the data points within a user-specifiable time
window centred on the current data point. Smoothing is usefully applied to the
output of the power program in order to damp ringing. A useful smoothing
window in this situation is one fourth the length of the Fourier transform
window. The rectify program replaces each data point with its absolute
value. This is useful for examining signals such as difference waves, in which
the important feature is the degree of deviation from zero. The range
program replaces each data point with the local range of the signal within a
user-specifiable time window centred on the current data point. This is a
measure of local amplitude at frequencies whose periods are comparable to the
length of the window.
Statistical analyses. In what follows, I use the ideas of data
matrices and probability matrices. A data matrix is the structure quite
familiar to all evoked potential researchers: it's what I was referring to
above as an averaged data file. If the sampling epoch contains n data
points, and c channels are being recorded, the result of averaging the
electrophysiological responses to each target event is a cxn
matrix of potentials. Thus the data matrix describes the actual
perturbations of the electroencephalogram over time. These perturbations are
superpositions of experimental effects and noise, and it isn't explicit from
the data matrix alone which is which. In order to represent the levels of
statistical significance of perturbations in the electroencephalogram,
Gnuroscan uses a probability matrix. The probability matrix is another
cxn matrix which maps strightforwardly onto the data matrix.
Instead of potentials, though, the entries in the probability matrix are levels
of statistical significance. These probabilities may be calculated by any of
Gnuroscan's several statistical programs. Probability matrices are stored in
the same format as data matrices and therefore may be viewed and manipulated
with any of the tools that view and manipulate data matrices. In particular,
applications needing a Bonferroni correction for multiple comparisons may
implement this by applying the scale program to the probability
matrix.
The ttest program implements a pointwise Student's t test on a
set of data matrices, and produces a probability matrix. The inputs are
treated as difference waves, and tested for significant deviation from zero.
This simple test is useful when examining a single hypothesis or comparison.
The chisquare program operates on z-transformed data matrices.
It uses a  2 test to determine whether the set of
z-scores differs significantly from samples that might be expected from
the standard normal distribution. A significant probability value indicates
that the stimulus does perturb the EEG, but not necessarily that it perturbs
the EEG in a consistent manner: a stimulus that causes an amplitude increase in
half the subjects and an amplitude decrease in the other half, for example,
would still produce a significant 2 probability even though the
mean post-stimulus amplitude would still be within the pre-stimulus baseline
range. This is a useful test when one is comparing only two treatments or
conditions. As a preprocessing step to this procedure, the ztrans
program transforms a data matrix to z-scores with respect to the
mean and variance of a baseline data matrix. Depending on the design of the
experiment, the baseline may be a subinterval of the epoch under consideration,
or it may be a separate average. A baseline interval may be extracted from a
longer epoch by means of the subepoch program.
2 test to determine whether the set of
z-scores differs significantly from samples that might be expected from
the standard normal distribution. A significant probability value indicates
that the stimulus does perturb the EEG, but not necessarily that it perturbs
the EEG in a consistent manner: a stimulus that causes an amplitude increase in
half the subjects and an amplitude decrease in the other half, for example,
would still produce a significant 2 probability even though the
mean post-stimulus amplitude would still be within the pre-stimulus baseline
range. This is a useful test when one is comparing only two treatments or
conditions. As a preprocessing step to this procedure, the ztrans
program transforms a data matrix to z-scores with respect to the
mean and variance of a baseline data matrix. Depending on the design of the
experiment, the baseline may be a subinterval of the epoch under consideration,
or it may be a separate average. A baseline interval may be extracted from a
longer epoch by means of the subepoch program.
The 2 test is fine for a first cut, but it has two major
shortcomings: by itself it fails to differentiate between differences in
variance (inconsistent perturbations) and differences in mean (consistent
perturbations), and it assumes that the distribution of individual amplitude
perturbations is normal. A more versatile test, given the speed of today's
electronic computers, is a permutation test. Often one desires to check whether
the event-related potentials or perturbations of one experimental group differ
from another, or from a control group. The usual way of accomplishing this is
Fisher's F-test. Nonparametric methods, however, are more accurate and
often more powerful than such resorts, because they have no need for an
assumption of normality. The permutation test is the most straightforward of
these. It compares the mean of the experimental group to the means of all
possible samples drawn from the pooled groups that are of the same size as the
experimental group. The ratio of the number of means less eccentric than the
experimental mean to the total number of means is the probability of the null
hypothesis. Gnuroscan implements a pointwise, exhaustive permutation test (the
permtest program), using Chase's [1970] algorithm for generating all
possible combinations of a given size. For cases in which the number of
possible combinations is prohibitively large, a Monte Carlo version of this
test also is available. (In the default configuration this critical value is
set to 105 combinations, but it can be tuned according to the speed
of floating-point computations on the implementation system.)
Another useful nonparametric test is the sign test, implemented by the
signtest program. This procedure applies the binomial probabilities to
the sign of the signal. It's most usefully applied to difference waves between
two experimental conditions, where, under the null hypothesis, positive and
negative differences are equally likely at any given point in the epoch. A
useful preprocessing step for this pointwise sign test is the transformation of
the difference wave with the extreme program, which filters the input
such that each point in the epoch is replaced by the point within a local
window whose absolute value is largest. When the width of the local window is
the period of the SSEP, this transformation has the effect of lining up the
peak amplitudes across all the subjects, thus increasing the power of this
pointwise test. Assuming that noise is symmetrically distributed with zero
mean--a fairly safe assumption in a difference wave--the probability of
positive sign remains one half even after this transformation, and therefore
the sign test still applies.
The mannwhitney program, like the permtest program, compares two
groups of data matrices. A pointwise Mann-Whitney U test is used. The
current implementation does not produce a probability matrix automatically, but
such a matrix may be constructed from the matrix of U values that is
output, using a standard statistical table. Applications of this program
include comparison of a control group and an experimental group, and also
comparisons of two treatment conditions within a single group of subjects.
The friedman program applies Friedman's test for a randomised block
design to a group of subjects each of whom receives each of several
experimental treatments. (Thus the individual subjects are the `blocks'.) This
is useful in cases in which there are several levels of the experimental
condition--for example, stimulus onset ansynchrony in a Posner-style attention
task--and each of those conditions is applied to each subject. The
mannwhitney program may be used as a pairwise post hoc test in
cases in which Friedman's test indicates significant variation.
Visualisation. Gnuroscan provides three visualisation routines,
all of which share characteristics. Each of these plotting programs produces a
PostScript program from a group of data matrices. The image coded by this
output program respresents variations in the signal and also the significance
levels of those variations.
plotavg uses Efron's [1982] bootstrap procedure to construct a
distribution for each channel from points in a baseline immediately prior to
the zero time point of the epoch. The bootstrap estimates the shape of the
distribution by repeatedly calculating means of subsets of the data points
drawn with replacement. This resampling procedure is computationally more
intensive than parametric procedures, but, assuming sufficient length of the
baseline interval from which the distribution is constructed, it yields a more
accurate representation of the actual distribution. The length of the baseline
may be tuned according to the needs of the particular application. Significant
(p<0.05, two-tailed) deviations from this bootstrapped baseline
distribution are plotted in shades of dark grey whose intensities correspond to
the significance level. Non-significant deviations are plotted in light
grey.
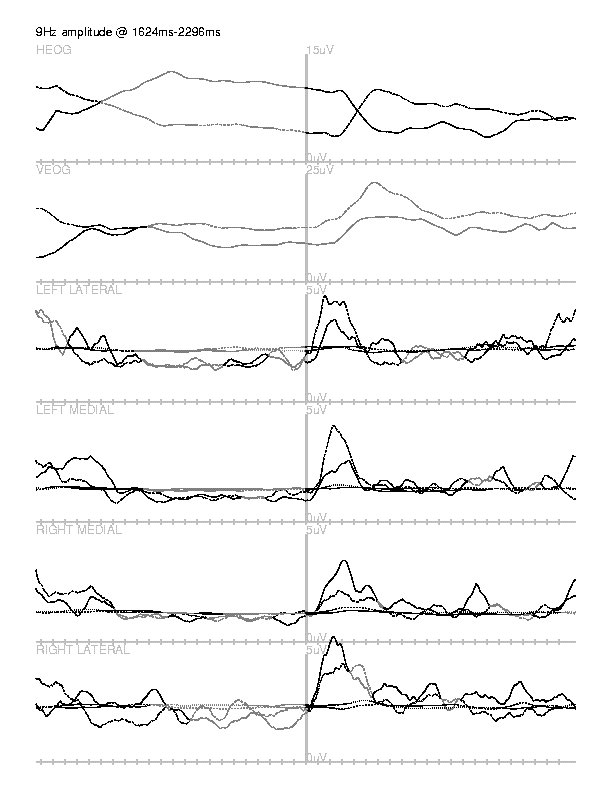 fplotavg also uses a nonparametric, combinational statistical test, but
its measure is the integral of a difference wave rather than the magnitude of
the current sample. Its parameters are pairs of data matrices. For each pair,
the difference wave is computed. The measure of difference is the integral
between zero-crossings of this difference wave, in each channel. In order to
estimate the magnitudes of such integrals due to random fluctuations, the
prestimulus baselines are slid along each other in a Monte Carlo manner and the
magnitudes of the integrals between zero-crossings of the resulting difference
wave are computed in order to establish an empirical distribution. Poststimulus
integrals are then ranked within this distribution in order to ascertain their
significance.
fplotavg also uses a nonparametric, combinational statistical test, but
its measure is the integral of a difference wave rather than the magnitude of
the current sample. Its parameters are pairs of data matrices. For each pair,
the difference wave is computed. The measure of difference is the integral
between zero-crossings of this difference wave, in each channel. In order to
estimate the magnitudes of such integrals due to random fluctuations, the
prestimulus baselines are slid along each other in a Monte Carlo manner and the
magnitudes of the integrals between zero-crossings of the resulting difference
wave are computed in order to establish an empirical distribution. Poststimulus
integrals are then ranked within this distribution in order to ascertain their
significance.
zplotavg plots a single data matrix. Like the other plotting programs,
it codes the significance level of each point as a grey level in the image.
Unlike these other programs, though, it reads the significance levels from an
accompanying probability matrix rather than computing them itself. Thus
zplotavg is quite useful in combination with output from the separate
statistical programs mentioned above.
STATUS REPORT
Taken together, these utilities form a flexible toolkit for the
manipulation of averaged electroencephalographic data. The scoring and
averaging routines are specific to the SSEP paradigm, but the other
routines--transformations, statistical tests, and visualisation programs--are
applicable to any problem in biological signal analysis.
The Gnuroscan system has been applied in a study of visual spatial attention
[Belmonte 1998]. This study found significant perturbations of both
phase-locked and non-phase-locked amplitude in various time intervals following
the presentation of a stimulus that cued a voluntary shift of visual spatial
attention. We hypothesise that these perturbations reflect the
neural processes underlying orienting to a stimulus, reorienting to a new
location, and distributing attention in anticipation of a subsequent shift.
Although the effects of static allocation of visual spatial attention have been
investigated using event-related potentials, this is the first study to apply
detailed ERP measures to rapidly repetitive shifts in visual spatial
attention.
LESSONS LEARNT
Several innovations implemented in the Gnuroscan system have been
valuable in analysis of steady-state evoked potentials and have avoided the
pitfalls created by the rapid stimulus delivery which is essential in
steady-state paradigms. Probabilistic constraints on pseudorandom stimulus
sequencing enable unambiguous mapping of behavioural responses to particular
target stimuli, even when the stimulus presentation rate is quite high.
Redundancy in coding of behavioural responses allows reconstruction of
behavioural data that are lost during rapid transfers. The use of
variable-length epochs in the averaging routine makes maximal use of the
steady-state response recorded between each pair of target stimuli, and the
inhomogeneity of sampling noise introduced by this feature can be compensated
for.
In addition, several of Gnuroscan's features have impact beyond the scope of
SSEPs. Fast digital filters automate the detection of saccades and blinks that
may be produced in response to rapid stimuli. Separate analyses of
phase-locked and non-phase-locked responses yield complementary information
about the neurophysiological processes associated with stimulus perception.
Nonparametric statistical tests, some of which can involve very substantial
amounts of computation, can give more exact and valid information on the
significance of the effects observed.
FUTURE PLANS
The focus in this work has been on development of the underlying
algorithms. Gnuroscan's user interface, therefore, is minimal, and is entirely
text-based. A priority in future development work will be to add a versatile,
graphical front end as a user interface to all of Gnuroscan's subprograms. In
addition, many statistical routines remain to be implemented; development of
the existing set of statistical programs was driven by immediate need for them
in a research application. The possibility exists for the development of an
interface to a commercial statistical package. Finally, in order to see the
widest possible application, future versions of Gnuroscan will have to include
translators that convert between the SCAN format and other major formats for
the storage of digital electroencephalographic data.
ACKNOWLEDGEMENTS
I thank Eric Courchesne of the Autism and Brain Development Research
Laboratory for providing facilities and financial support for part of this
work, and, more importantly, for his friendship which spanned most of my time
in San Diego. I also thank Brian Egaas of Science Applications International
Corporation for teaching me the finer points of signal processing, Bruce
Fischer of the MIT Mathematics Department for suggesting conditioned
distributions as a way to implement constraints on pseudorandom inter-target
intervals, Anthony Gamst of the UCSD Mathematics Department for suggesting a
more powerful, computational alternative to parametric statistics for the
analysis of moving-window amplitudes, and Robert Ringrose of the MIT Artificial
Intelligence Laboratory for providing computing facilities for the latter part
of this work.
BIBLIOGRAPHY
Natacha Akshoomoff, Eric Courchesne, `A New Role for the Cerebellum in
Cognitive Operations', Behavioral Neuroscience 106:5:731-738 (1992).
D Allen, AM Norcia, CW Tyler, `Comparative Study of Electrophysiological and
Psychophysical Measurement of the Contrast Sensitivity Function in Humans',
American Journal of Optometry and Physiological Optics 63:6:442-449
(1986).
M Bach, T Meigen, `Electrophysiological Correlates of Texture Segregation in
the Human Visual Evoked Potential', Vision Research 32:3:417-424
(1992).
Matthew Belmonte, `Shifts of Visual Spatial Attention Modulate a Steady-State
Visual Evoked Potential', Cognitive Brain Research 6:4:295-307 (1998).
Phillip J Chase, `Algorithm 382: Combinations of M out of N
Objects [G6]', Communications of the Association for Computing Machinery
13:6:368 (1970).
SA Chen, LZ Wu, DZ Wu, `Objective Measurement of Contrast Sensitivity Using the
Steady-State Visual Evoked Potential', Documenta Ophthalmologica
75:2:145-153 (1990).
Eric Courchesne, Jeanne Townsend, Natacha Akshoomoff, Rachel Yeung-Courchesne,
Alan Lincoln, Gary Press, James Murakami, Hector James, Osamu Saitoh, Brian
Egaas, Richard Haas, Laura Schreibman, `A New Finding: Impairment in Shifting
Attention in Autistic and Cerebellar Patients'. In: SH Broman & J Grafman
(eds.), Atypical Cognitive Deficits in Developmental Disorders: Implications
for Brain Function (Hillsdale, New Jersey: Lawrence Erlbaum, 1994a),
101-137.
Eric Courchesne, Jeanne Townsend, Natacha Akshoomoff, Osamu Saitoh, Rachel
Yeung-Courchesne, Alan Lincoln, Hector James, Richard Haas; Laura Schreibman,
Lily Lau, `Impairment in Shifting Attention in Autistic and Cerebellar
Patients', Behavioral Neuroscience 108:5:848-865 (1994b).
Bradley Efron, The Jackknife, the Bootstrap and Other Resampling Plans.
Philadelphia: Society for Industrial and Applied Mathematics, 1982.
Alexander I Fedotchev, Alexander T Bondar, Vladimir F Konovalov, `Stability of
resonance EEG reactions to flickering light in humans', International
Journal of Psychophysiology 9:2:189-193 (1990).
Robert Galambos, Scott Makeig, `Physiological Studies of Central Masking in
Man. I: The Effects of Noise on the 40-Hz Steady-State Response', Journal of
the Acoustical Society of America 92:5:2683-2690 (1992).
Robert Galambos, Scott Makeig, PJ Talmachoff, `A 40-Hz Auditory Potential
Recorded from the Human Scalp', Proceedings of the National Academy of
Sciences 78:4:2643-2647 (1981).
Bo Hjorth, `An On-Line Transformation of EEG Scalp Potentials into Orthogonal
Source Derivations', Electroencephalography and Clinical Neurophysiology
39:5:526-530 (1975).
William H Press, Saul A Teukolsky, William T Vetterling, Brian P Flannery,
Numerical Recipes in C: the art of scientific computing 2/e. New York:
Cambridge University Press, 1992.
JW Rohrbaugh, JL Varner, SR Paige, MJ Eckardt, RJ Ellingson, `Event-Related
Perturbations in an Electrophysiological Measure of Auditory Function: a
Measure of Sensitivity during Orienting?', Biological Psychology
29:3:247-271 (1989).
Richard B Silberstein, Mark A Schier, Andrew Pipingas, Joseph Ciorciari,
Stephen R Wood, David G Simpson, `Steady-State Visually Evoked Potential
Topography Associated with a Visual Vigilance Task', Brain Topography
3:2:337-347 (1990).
Richard B Silberstein, Joseph Ciorciari, Andrew Pipingas, `Steady-state
visually evoked potential topography during the Wisconsin card sorting test',
Electroencephalography and Clinical Neurophysiology 96:1:24-35 (1995).
RJ Snowden, D Ullrich, M Bach, `Isolation and Characteristics of a Steady-State
Visually-Evoked Potential in Humans Related to the Motion of a Stimulus',
Vision Research 35:10:1365-1373 (1995).